Doing different things with the two hands at the same time, like rubbing your stomach while padding your head, is difficult. Drawing a circle with the left hand while drawing a square with the right hand is nearly impossible. Studying interference between the hands illuminates the "dark side" of bimanual coordination. While the two hands normally benefit from "talking" to each other to achieve one common goal, this cross-talk is detrimental when we want each hands to perform independent actions. By finding the mechanisms through which the hands interfere with each other when they move independently, researchers hope to learn how the hands interact when they have to move coordinated.
One task that had been used extensively to study bimanual interference is continuous circle drawing. You can perform the self-experiment by drawing circles with both hands at a pretty high speed. Most likely you will prefer to draw circles in a mirror-symmetric fashion, i.e. one hand moves clockwise, the other counter-clockwise. The spatial quality of circles drawn symmetrically is higher than those drawn asymmetrically, with both hands moving clockwise or counter-clockwise (Semjen et al., 1995). Also, when you try to increase the speed of asymmetric movements and go as fast as you possibly can, you will most likely show some spontaneous transitions into symmetric performance (Kelso, 1984). These all are indications for your natural preference to move the two upper limbs in mirror symmetry.
Interactions on the spinal level
One hypothesis is that these interactions are happening very close to the motor output, i.e. on the level of the spinal cord. The idea here is that phylogenic older behaviors, such as swimming or walking, have put into place the preference for in-phase or anti-phase movements. While most of the fibers from the left motor cortex go to the right hand, about 10% of all cortico-spinal fibers go uncrossed to the left hand. It is possible that these projections make it easier to perform mirror-symmetric movements (Carson, 2005). An extreme example of such cross-talk can be observed in people who are born with the "congenital mirror syndrome". These individuals probably have many more uncrossed fibers, and when they perform an actions with one hand, the other hand shows small involuntary twitches.
However, it is unlikely that the bimanual interference in normal individuals is caused by these fibers. This conclusion can be drawn from the study of patients who underwent a surgical removal of the corpus callosum, a massive fiber bundle connecting the two hemispheres. These patients are actually able to draw a circle and a square at the same time (Franz et al. 1996). In the bimanual circle drawing task, callosotomy patients do not show a preference for symmetric movements (Kennerley et al., 2003). They exhibit something much stranger: one hand can move at a different speed than the other hand, so that the hands lap each other. This shows that fibers connecting the two hemisphere, rather than uncrossed cortical-spinal fibers, are the main cause for the tendency to move the upper limbs in symmetry.
Interactions of movement parameters or goal representations?
In a series of studies, Herbert Heuer and colleagues (e.g., Heuer, 1993) have shown that many interference effects are "transient", that is they disappear when subjects have enough time to mentally prepare their movements . Such findings have led to the hypothesis that cross-talk arises through interactions during movement planning, that is, when the parameters of the movements are specified. In research conducted at the Cognition and Action Lab at University of California, Berkeley, we have focused on the notion that interference between bimanual actions can depend heavily on the way the movements are instructed, and consequently, represented by the participant. For example, the time to initiate movements with different movement parameters is greater than the time to initiate symmetric movements, but only when the movement are instructed with symbolic cues. When movements are specified by presenting the target locations directly, the movement parameters are not part of the action representation, and consequently no interference between the two actions arises (Diedrichsen et al., 2001).
 |
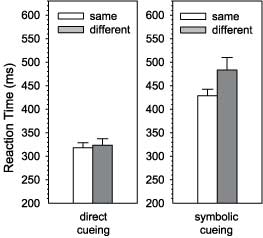 |
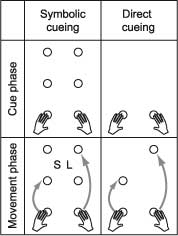
Figure 1. The selection of bimanual movements can either be a direct consequence of the presentation of the targets (direct cueing), or can be based on symbolic cues indicating a movement parameter (symbolic cueing). Here the indicated parameter is movement amplitude with S for a short and L for a long movements. |
Figure 2. The reaction time cost in the initiation of bimanual movements is restricted to a situation, in which movements are cued symbolically. When cued directly, movement initiation does not show any cost of incongruent movements (short-long). This holds true even compared to a situation in which only one of the movements has to be initiated (Diedrichsen et al., 2001). |
Movements of different amplitudes can sometimes be initiated even more quickly than movements of same amplitudes. This is the case, when the movements of different amplitude are directed to targets of the same color, and the movements of the same amplitude are directed to targets of different colors (Diedrichsen et al., 2003a).
Thus, we think that much of bimanual interference is based on a representation of abstract actions codes. These action codes may encompass the features of the targets of reaching movements or the intended consequences of actions (Hommel, 2001). This interference can be found to be abolished following callosotomy (the surgical removal of the fiber-band that connects the two cerebral hemispheres). For example, when callosotomy patients have to produce short force pulses with their left and right hand, they are as fast to initiate force pulses of the same as different target amplitudes. In contrast, age-matched controls show a substantial reaction time cost in this situation (Diedrichsen et al., 2003b).
In agreement with this finding, Mechsner et al. (2001) recently reported that bimanual interactions in the continuous circle drawing task can be dramatically altered when the perceptual consequences of such movements are manipulated. Both findings argue that the interference in this bimanual task is based rather on a more abstract representation of the action than on low-level interactions between the motor implementation processes of the two hands.
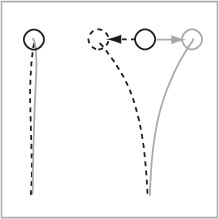 Another dramatic demonstration of the independence of the two hands comes from a study using online adjustments during bimanual reaching movements
(Diedrichsen et al. 2004).
Adjustments of hand trajectory occur very quickly (under 200 ms) when a target is displaced during a reaching movement.
In the bimanual case, even when both targets are displaced in different
directions, these rapid corrections are also observed and only delayed by 20 ms compared to unimanual reaches, with little change observed in movement
velocity and accuracy. These results suggest two independent "autopilots", ensuring that each
hand reaches its target location. These corrections do not require eye movements or shifts of
visual attention towards the displaced target(s). However, when one hand adjusts
for a target displacement to the left (dashed) or to the right (gray), the other
hand is briefly perturbed in the same direction (see Figure). While the source
of this cross-talk remains to be determined, the main insight is that the
interference in bimanual movements is strongly reduced or absent when we
directly reach for targets or when other external stimuli help us represent
these otherwise difficult movements. Thus, it is most likely the difficulty to
represent two conflicting spatial plans that limits our ability to pad our head
while rubbing our stomach. Put simply, we shouldn't blame the hands for a
limitation in our heads.
Another dramatic demonstration of the independence of the two hands comes from a study using online adjustments during bimanual reaching movements
(Diedrichsen et al. 2004).
Adjustments of hand trajectory occur very quickly (under 200 ms) when a target is displaced during a reaching movement.
In the bimanual case, even when both targets are displaced in different
directions, these rapid corrections are also observed and only delayed by 20 ms compared to unimanual reaches, with little change observed in movement
velocity and accuracy. These results suggest two independent "autopilots", ensuring that each
hand reaches its target location. These corrections do not require eye movements or shifts of
visual attention towards the displaced target(s). However, when one hand adjusts
for a target displacement to the left (dashed) or to the right (gray), the other
hand is briefly perturbed in the same direction (see Figure). While the source
of this cross-talk remains to be determined, the main insight is that the
interference in bimanual movements is strongly reduced or absent when we
directly reach for targets or when other external stimuli help us represent
these otherwise difficult movements. Thus, it is most likely the difficulty to
represent two conflicting spatial plans that limits our ability to pad our head
while rubbing our stomach. Put simply, we shouldn't blame the hands for a
limitation in our heads.
Neural mechanisms of interference
In recent work (Diedrichsen, in press) we studied neural correlated of interference in
the bimanual reaching task in functional magnetic resonance imaging (fMRI). Participants executed
symmetric or asymmetric reaching movements either cued by the spatially by
presenting the targets directly, or symbolically by presenting letters
(F for forward, S for sideways). A large reaction time cost was again only observed when symbolic cues
were in conflict (respond to FS, compared to FF). This conflict caused
increased activity in PreSMA and the cingulate gyrus, the same areas that are
active when you try to perform cognitive interference tasks, such as the
Stroop-test.
This supports our notion that bimanual interference occurs when we try to decide
what to do with each hand, rather than how to do it.
However, both in the symbolic and spatial cueing conditions, we observed
activity in the posterior aspect of the superior parietal lobe, an area that is
responsible for spatial planning of movements. Even though participants
can perform bimanual movements in asymmetric directions rather effortlessly, the
interference occurring during the spatial planning of the movement left a neural
trace.
References
- Carson, R. G. (2005). Neural pathways mediating bilateral interactions between the upper limbs. Brain Research. Brain Research Reviews, 49(3), 641-662.
- Diedrichsen, J., Grafton,
S., Albert, N., Hazeltine, E., & Ivry, R. B. (in press). Goal-selection and
movement-related conflict during bimanual reaching movements. Cerebral Cortex.

- Diedrichsen, J., Hazeltine, E., Kennerley, S., Ivry, R. B. (2001).
Moving to directly cued locations abolishes spatial interference during bimanual actions.
Psychological Science, 12, 493-498.
- Diedrichsen, J., & Hazeltine, E. (2001). Unifying by binding: will binding really bind? A commentary on Hommel et al. (2001).
Behavioral & Brain Sciences, 24, 884-885.
- Diedrichsen, J., Ivry, R. B., Hazeltine, E., Kennerley, S., & Cohen, A. (2003a).
Bimanual interference associated with the selection of target locations.
Journal of Experimental Psychology: Human Perception & Performance, 29(1), 64-77.
- Diedrichsen, J., Hazeltine, E., Nurss, W., & Ivry, R. B. (2003b).
The role of the corpus callosum in the coupling of
bimanual isometric force pulses. Journal of Neurophysiology, 90(4), 2409-2418.
- Diedrichsen, J., Nambisan, R., Kennerley, S., & Ivry, R. B. (2004). Independent on-line control of
the two hands during bimanual reaching. European Journal of Neuroscience, 19, 1643-1652.
- Hazeltine, E., Diedrichsen, J., Kennerley, S., & Ivry, R. B. (2003).
Bimanual cross-talk during reaching movements is primarily related to response
selection, not the specification of motor parameters.
Psychological Research, 67, 56-70.
- Heuer, H. (1993). Structural constraints on bimanual movements. Psychological Research/Psychologische Forschung, 55(2), 83-98.
- Hommel, B., M�sseler, J., Aschersleben, G., & Prinz, W. (2001). The theory of event coding (TEC): A framework for perception and action planning. Behavioral and Brain Sciences, 24(4).
- Ivry, R. B., Diedrichsen, J., Spencer, R. M., Hazeline, E., & Semjen, A. (2004). A cognitive neuroscience perspective on bimanual coordination and interference. In S. Swinnen & J. Duysens (Eds.), Interlimb Coordination (pp. 259-295). Boston: Kluwer Academic Publishing.
- Kennerley, S., Diedrichsen, J., Hazeltine, E., Semjen, A., & Ivry, R. B. (2002).
Callosotomy patients exhibit temporal and spatial uncoupling during continuous bimanual movements.
Nature Neuroscience, 5(4), 376-381.
- Kelso, J. A. S. (1984). Phase transitions and critical behavior in human bimanual coordination. Am J Physiol, 246(6 Pt 2), R1000-1004.
- Mechsner, F., Kerzel, D., Knoblich, G., & Prinz, W. (2001). Perceptual basis of bimanual coordination. Nature, 414(6859), 69-73.
- Semjen, A., Summers, J. J., & Cattaert, D. (1995). Hand coordination in bimanual circle drawing. Journal of Experimental Psychology: Human Perception & Performance, 21(5), 1139-1157.